




还剩52页未读,
继续阅读
第06章 蛋白质分选与膜泡运输课件PPT
展开
这是一份第06章 蛋白质分选与膜泡运输课件PPT,共60页。PPT课件主要包含了本章主要内容,细胞内蛋白质的分选,细胞内膜泡运输,第一节,第二节等内容,欢迎下载使用。
信号假说与蛋白质分选信号
蛋白质分选的基本途径与类型
蛋白质向线粒体和叶绿体的分选
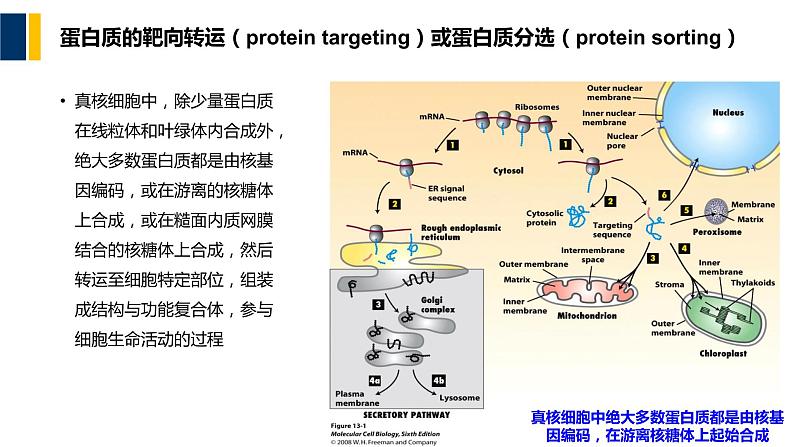
真核细胞中绝大多数蛋白质都是由核基因编码,在游离核糖体上起始合成
蛋白质的靶向转运(prtein targeting)或蛋白质分选(prtein srting)
真核细胞中,除少量蛋白质在线粒体和叶绿体内合成外,绝大多数蛋白质都是由核基因编码,或在游离的核糖体上合成,或在糙面内质网膜结合的核糖体上合成,然后转运至细胞特定部位,组装成结构与功能复合体,参与细胞生命活动的过程
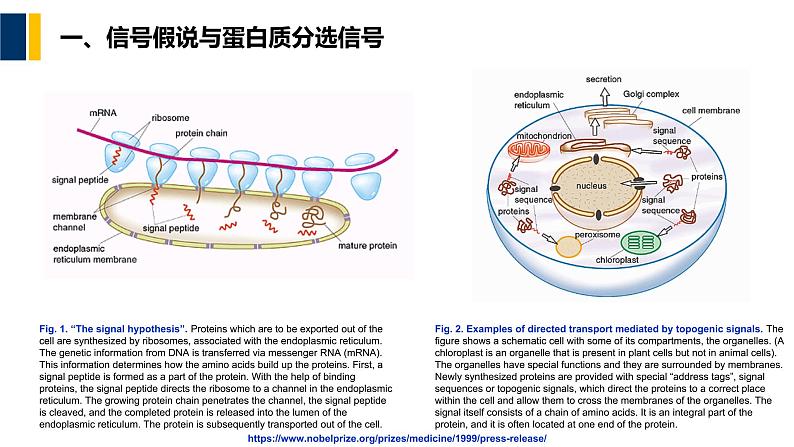
一、信号假说与蛋白质分选信号
Fig. 2. Examples f directed transprt mediated by tpgenic signals. The figure shws a schematic cell with sme f its cmpartments, the rganelles. (A chlrplast is an rganelle that is present in plant cells but nt in animal cells). The rganelles have special functins and they are surrunded by membranes. Newly synthesized prteins are prvided with special “address tags”, signal sequences r tpgenic signals, which direct the prteins t a crrect place within the cell and allw them t crss the membranes f the rganelles. The signal itself cnsists f a chain f amin acids. It is an integral part f the prtein, and it is ften lcated at ne end f the prtein.
Fig. 1. “The signal hypthesis”. Prteins which are t be exprted ut f the cell are synthesized by ribsmes, assciated with the endplasmic reticulum. The genetic infrmatin frm DNA is transferred via messenger RNA (mRNA). This infrmatin determines hw the amin acids build up the prteins. First, a signal peptide is frmed as a part f the prtein. With the help f binding prteins, the signal peptide directs the ribsme t a channel in the endplasmic reticulum. The grwing prtein chain penetrates the channel, the signal peptide is cleaved, and the cmpleted prtein is released int the lumen f the endplasmic reticulum. The prtein is subsequently transprted ut f the cell.
The Nbel Prize in Physilgy r Medicine 1999
"fr the discvery that prteins have intrinsic signals that gvern their transprt and lcalizatin in the cell."
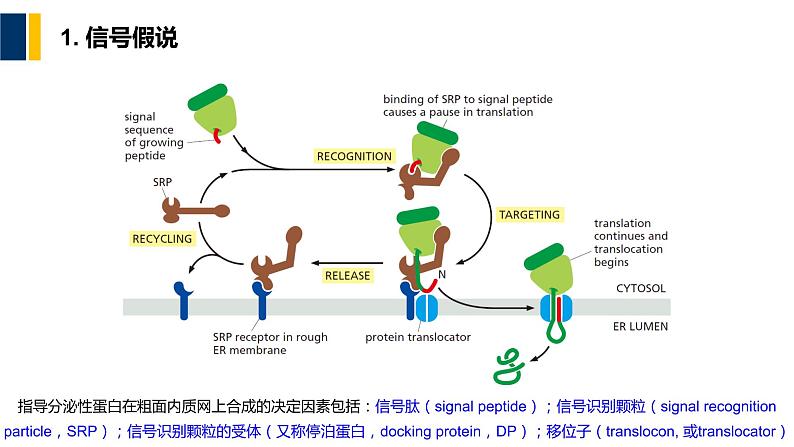
指导分泌性蛋白在粗面内质网上合成的决定因素包括:信号肽(signal peptide);信号识别颗粒(signal recgnitin particle,SRP);信号识别颗粒的受体(又称停泊蛋白,dcking prtein,DP);移位子(translcn, 或translcatr)
(1)信号肽(signal peptide)
位于蛋白质的N 端,一般由16~26 个残基组成;似乎没有严格的专一性包括疏水核心区、信号肽的C-端和N-端等3 部分原核细胞某些分泌性蛋白的N 端也具有信号序列
图6-1 信号肽的一级结构序列以血清白蛋白和HIV-1 型病毒的糖蛋白gp160 信号肽为例,显示出两者信号肽一级序列分别由疏水核心、C- 端和N- 端三个区域构成
(2)信号识别颗粒(SRP)
由6 种不同的蛋白质和一个由300 个核苷酸组成的 7S RNA 结合组成的一种核糖核蛋白复合体通常存在于细胞质基质中;既可与新生肽信号序列和核糖体大亚基结合,又可与内质网膜上SRP 受体结合
图6-2 信号识别颗粒(SRP)的结构示意图含有两个结构域,P54 蛋白是一种包含成簇甲硫氨酸残基的GTP酶,甲硫氨酸侧链与信号肽的疏水核心结合;当SRP 与信号肽结合后,SRP 的P9 和P14 蛋白复合体阻断新生肽链的翻译
(3)SRP 受体(Signal Recgnitin Particle Receptr)
内质网膜上的整合蛋白,由α 和β 亚基组成,可特异地与SRP 结合SRP 的p54 亚基和SRP 受体的α 亚基与GTP 结合时,会增进SRP/ 新生肽/ 核糖体复合物与SRP 受体结合的强度。
Dynamic behavir f SRP upn SR interactin
DOI: 10.1126/science.1124864
体外非细胞系统(cell free system)进行蛋白质合成实验,证实分泌性蛋白向rER(micrsme,微粒体)腔内的转运是同蛋白质翻译过程偶联进行的,这种分泌蛋白在信号肽引导下边翻译边跨膜转运的过程称为共翻译转运 (ctranslatinal translcatin)
蛋白质翻译过程与SRP、DP和微粒体的关系
在非细胞系统中蛋白质的翻译过程与SRP、DP 和微粒体的关系
* “+”和“-”分别代表反应混合物中存在(+)或不存在(-)该物质
(4)分泌性蛋白的合成与共翻译转运过程
图6-3 分泌性蛋白的合成与共翻译转运过程图解
分泌性蛋白的合成与其共翻译转运
Sec61 Prtein Translcatr
(5)内质网膜整合蛋白的信号序列
起始转移序列(start transfer sequence)内在停止转移锚定序列(internal stp-transfer anchr sequence, STA)内在信号锚定序列(internal signal anchr sequence, SA)
内质网膜整合蛋白的拓扑学类型
图6-4 内质网膜整合蛋白的拓扑学类型
单次跨膜蛋白插入转移机制:新生跨膜蛋白肽链中既有N端信号肽,又有停止转移信号(stp transfer signal)
整合膜蛋白的合成——单次跨膜
单次跨膜蛋白的插入转移机制:新生跨膜蛋白的肽链中没有N 端信号肽,只有内部信号序列(internal signal sequence)。内部信号序列的转位方向主要取决于其侧翼氨基酸残基电荷的分布。一般而言,带正电荷的氨基酸残基一侧朝向细胞质基质一侧
多次跨膜蛋白的插入转移机制:多次跨膜蛋白肽链中,常常有两个或两个以上的疏水性开始转移信号序列和停止转移信号序列。一般认为,多次跨膜蛋白是以内部信号序列(internal signal sequence)作为其开始转移信号
整合膜蛋白的合成——多次跨膜
蛋白质自身所携带的分选信号需要被互补的分选受体所识别,然后在分选受体帮助下,引导蛋白质转运到目的地
2. 蛋白质分选信号序列
导肽(leader peptide):指导在细胞质基质中合成的蛋白质进入线粒体、叶绿体及过氧化物酶体的信号序列
表6-2 指导蛋白质从细胞质基质转运到细胞器的靶向序列的主要特征
2. 蛋白质分选信号序列(signal sequence)
两类分选信号一类为连续的氨基酸序列,一般长15~60个氨基酸残基。完成分选过程后,有些这类信号序列会被特异性酶从成熟蛋白质上切除一类为三维结构的信号斑,在蛋白折叠完成后,其表面原子按一定的间结构排布形成。形成信号斑的氨基酸残基可能在线性氨基酸序列上相距很远。信号斑会保留在成熟蛋白质中不被切除
途径(1)翻译后转运途径(2)共翻译转运途径类型(1)蛋白质的跨膜转运(transmembrane transprt)(2)膜泡运输(vesicular transprt)(3)选择性的门控转运(gated transprt)(4)细胞质基质中蛋白质的转运
二、蛋白质分选转运的基本途径与类型
真核细胞蛋白质分选的主要途径与类型
图6-5 真核细胞蛋白质分选的主要途径与类型
三、蛋白质向线粒体和叶绿体的分选
转运到线粒体、叶绿体和过氧化物酶体等中的蛋白需要多个不同靶向序列(targeting sequences);细胞器蛋白最终是定位在不同膜上还是不同基质空间,除N- 端不同转运肽外,还需要其他空间定位信号序列通过翻译后转运途径进入线粒体、叶绿体和过氧化物酶体等细胞器的蛋白,必须在分子伴侣帮助下解折叠或维持非折叠状态,蛋白质输入这些细胞器通常是需要能量的过程
1. 蛋白质从细胞质基质输入到线粒体
蛋白质从细胞质基质输入到线粒体基质蛋白质以3 种途径从细胞质基质输入到线粒体内膜线粒体蛋白通过2 种途径从细胞质基质输入到线粒体膜间空间
图6-6 通过后翻译转运途径,核基因编码的线粒体蛋白的转运A. 蛋白质从细胞质基质输入到线粒体基质(图左上);B. 线粒体蛋白通过3 种途径从细胞质基质输入到线粒体内膜(图右);C. 线粒体蛋白通过2 种途径从细胞质基质输入到线粒体膜间空间(图左下)
外膜蛋白内膜蛋白叶绿体基质蛋白类囊体膜蛋白类囊体腔蛋白
2. 蛋白质从细胞质基质向叶绿体的分选
图6-7 通过后翻译转运途径,叶绿体蛋白从细胞质基质输入到类囊体腔注:Tm/Tim:Translcn f the uter/inner membrane
叶绿体蛋白从细胞质基质输入到类囊体腔
COP Ⅱ 包被膜泡的装配及运输
COP I 包被膜泡的装配与运输
网格蛋白/接头蛋白包被膜泡的装配与运输
转运膜泡与靶膜的锚定和融合
The Nbel Prize in Physilgy r Medicine 2013
"fr their discveries f machinery regulating vesicle traffic, a majr transprt system in ur cells."
James E. Rthman
James E. Rthman discvered that a prtein cmplex (pictured in range) enables vesicles t fuse with their target membranes. Prteins n the vesicle bind t specific cmplementary prteins n the target membrane, ensuring that the vesicle fuses at the right lcatin and that carg mlecules are delivered t the crrect destinatin.
Randy W. Schekman
Randy W. Schekman discvered genes encding prteins that are key regulatrs f vesicle traffic. Cmparing nrmal with genetically mutated yeast cells in which vesicle traffic was disturbed, he identified genes that cntrl transprt t different cmpartments and t the cell surface.
secretin-defective mutant strain (sec).
Thmas C. Südhf
Thmas C. Südhf studied hw signals are transmitted frm ne nerve cell t anther in the brain, and hw calcium cntrls this prcess. He identified mlecular machinery (pictured in purple) that senses calcium ins (Ca2+) and triggers vesicle fusin, thereby explaining hw tempral precisin is achieved and hw signaling substances can be released frm the vesicles n cmmand.
图6-8 蛋白质的分泌与胞吞途径概观1. 在rER 合成的蛋白质, 通过共翻译转运途径跨膜运输;2. 内质网出芽,形成转运泡并与高尔基体融合,形成cis- 高尔基网状结构;3. 从cis- 高尔基膜囊和cis- 高尔基体顺面网状结构到rER 的逆向运输;4. 高尔基膜囊从cis 面→ trans 面成熟递进(非膜泡过程);5. 从高尔基体后期膜囊→早期膜囊的逆向运输;6. 组成型分泌;7. 调节型分泌;8. 分选到溶酶体;9. 胞吞途径
Fig. A “rad-map” f the secretry and endcytic pathways
三种不同类型的包被膜泡
图6-9 在细胞合成- 分泌与内吞途径中三种不同的主要膜泡运输方式COP II(cat prtein II)包被泡、COPI(cat prtein I)包被泡和网格蛋白/ 接头蛋白(clathrin/adaptr prtein)包被泡,它们分别介导不同的膜泡转运途径
COPⅡ包被膜泡介导从内质网到高尔基体的顺向运输。COPⅠ包被膜泡介导细胞内膜泡逆向运输,负责从高尔基体反面膜囊到高尔基体顺面膜囊,以及从高尔基体CGN区到内质网的膜泡转运。网格蛋白/ 接头蛋白包被膜泡介导几种蛋白质分选途径,包括从高尔基体TGN向胞内体、溶酶体、黑(色)素体、血小板囊泡或植物细胞液泡的运输。另外,在受体介导的胞吞途径中还负责将物质从细胞表面运往胞内体转而到溶酶体的运输
蛋白质转运中涉及的3 种包被膜泡的特征比较
表8-3 蛋白质转运中涉及的三种包被膜泡的特征比较
COP Ⅱ 包被膜泡介导细胞内顺向运输(antergrade transprt),即负责从内质网到高尔基体的物质运输COP Ⅱ 包被蛋白组分小分子GTP 结合蛋白Sar1Sec23/Sec24 复合物Sec13/Sec31 复合物以及大的纤维蛋白Sec16
二、COP II包被膜泡的装配及运输
COP Ⅱ 包被膜泡的装配与去装配
图6-10 Sar1 蛋白在Cp Ⅱ 包被膜泡装配与去装配中作用模型1. Sar1 与膜结合,GTP 交换;2. COP Ⅱ包被装配;3. GTP 水解;4. COP Ⅱ包被去装配
Sar1 蛋白在CpⅡ包被膜泡装配与去装配中作用
细胞质中可溶性 Sar1- GDP 与ER 膜蛋白Sec12(鸟苷酸交换因子)相互作用, 催化GTP 置换GDP 形成Sar1-GTP,GTP 的结合引发 Sar1 构象改变暴露出疏水N 端并插入ER 膜,膜结合Sar1 对包被蛋白进一步装配起募集者作用
COP Ⅱ 包被膜泡装配
小分子GTP 结合蛋白Sar1
Sec23 是GTP酶活化蛋白(GTPase-activating prtein, GAP),能够提高Sar1水解GTP 的酶活性。特别是当Sec31 被招募到复合体中后,能够提高Sec23 作为Sar1的GAP 活性达10 倍,从而使成熟的COPII包被膜泡很容易发生去装配
COPⅡ包被膜泡的去装配
图6-11 不同类型的膜泡运输A. 内质网与高尔基体之间,分别由COPⅡ和COPⅠ膜泡介导蛋白质顺向和逆向转运:步骤1~3 表示由COPⅡ包被膜泡介导的顺向运输,内质网膜出芽,可溶性包被蛋白复合物聚合形成膜泡包被;v-SNAREs 和其它被转运的膜蛋白和可溶性蛋白(与芽泡膜上受体结合),通过与包被蛋白的相互作用被包装在膜泡内,包被复合物解聚再循环利用,v-SNAREs 蛋白在脱包被膜泡表面得以暴露,以利于同靶膜的融合。步骤4~6 表示由COPⅠ包被膜泡介导的反向运输,膜脂双分子层、某些蛋白如v-SNAREs 和错误分选的内质网驻留蛋白,从cis- 高尔基网状区转运到内质网。B. KDEL 受体在从高尔基体回收内质网腔驻留蛋白中的作用:步骤1~2 表示内质网腔中高水平存在的蛋白C- 端具有KDEL 信号,被包装到COPⅡ包被膜泡,并转运到高尔基体;步骤3~4 表示内质网逃逸蛋白通过COPⅠ膜泡介导的反向运输得以回收,注意,COPⅡ、COPⅠ和cis- 高尔基网状区膜上均有识别与结合KDEL 信号的受体,信号与受体的亲和力受到pH 高低的影响,低pH 值促进结合,高pH 值有利于释放。C. 在供体膜内质网出芽及其转运蛋白的包装示意图:通过募集小分子GTP 结合蛋白,引发供体膜出芽,然后,胞质中的包被蛋白复合物与被转运的膜蛋白胞质结构域结合,其中有些膜蛋白也作为腔内可溶性蛋白的受体
三、COP I 包被膜泡的装配及运输
COP I 包被膜泡介导细胞内膜泡逆向运输(retrgrade transprt),负责从高尔基体反面膜囊到高尔基体顺面膜囊以及从高尔基体顺面网状区到内质网的膜泡转运含有7 种不同的蛋白质亚基和一种调节膜泡转运的 GTP 结合蛋白ARF(ADP Ribsylatin Factr 1 )ARF 也是一种结合 GDP/GTP 转换的分子开关调控蛋白
Cat assembly drives vesicle frmatin
KDEL 受体主要定位在高尔基体TGN 区、COPⅡ和COPⅠ包被膜泡的膜上,它们能识别并结合KDEL 分选信号。如果在内质网发生错误包装和转运,由于COPⅠ膜泡上也有KDEL 受体,所以也能保证逃逸蛋白被内质网回收
COP Ⅱ和COP Ⅰ包被膜泡介导内质网与高尔基体之间蛋白质顺向和逆向转运
①~③表示由COPⅡ包被膜泡介导的顺向运输,内质网膜出芽,可溶性包被蛋白复合物聚合形成膜泡包被;v-SNARE,以及其他被转运的膜蛋白和可溶性蛋白(与芽泡膜上受体结合)通过与包被蛋白的相互作用被包装在膜泡内,包被复合物解聚再循环利用,v-SNARE蛋白在脱包被膜泡表面得以暴露,以利于同靶膜的融合。④~⑥ 表示由COPⅠ包被膜泡介导的逆向运输,膜脂双分子层、某些蛋白质(如v-SNARE和错误分选的内质网驻留蛋白)从高尔基体CGN区转运回内质网
网格蛋白/ 接头蛋白包被膜泡介导的蛋白质分选途径从高尔基体TGN向胞内体或向溶酶体、 黑(色)素体、血小板囊泡和液泡的运输受体介导的胞吞途径中负责将物质从细胞表面运往胞内体转而到溶酶体的运输
四、网格蛋白/接头蛋白包被膜泡的装配与运输
图6-12 网格蛋白、多角形网格包被结构及其的发动蛋白介导的网格蛋白/AP 包被小泡的形成示意图A. 三腿网格蛋白分子,3 条重链,3 条轻链;B. 在体外缺乏膜泡的情况下自组装的网格蛋白网格包被(基于PDB 数据库1XI4 结构绘制)。C. 发动蛋白介导的网格蛋白/AP 包被小泡的形成示意图
网格蛋白及其包被膜泡的形成
胞吞过程中网格蛋白/接头蛋白包被膜泡的装配
ARF 也参与网格蛋白/ 接头蛋白包被的起始装配(细胞膜—胞内体 不需要)发动蛋白(dynamin)具有GTPase 活性,导致网格蛋白/ 接头蛋白包被膜泡从供体膜断裂并释放
发动蛋白在网格蛋白包被膜泡形成中的作用
五、转运泡与靶膜的锚定与融合
① 供体膜的出芽、装配和断裂,形成不同的包被转运膜泡② 在细胞内由马达蛋白驱动、以微管为轨道的膜泡运输③ 转运膜泡与特定靶膜的锚定和融合
可溶性N-乙基马来酰亚胺敏感因子结合蛋白受体SNARE (sluble N ethyl maleimide-sensitive factr attachment prtein receptr)
Rab (ras-related in brain)
Ras (rat sarcma, Ras)
v-SNARE (vesicle SNARE)
t-SNARE (target SNARE)
1. Rab蛋白参与转运膜泡与靶膜的识别与锚定
胞质中Rab 蛋白在特异性鸟苷酸交换因子(GEF)催化下,Rab-GDP 转换为Rab-GTP,构象改变致使其通过类异戊二烯基团插入转运膜泡表面,与靶膜上Rab 效应器结合蛋白相互作用,从而使转运膜泡被锚定在靶膜上
Rab 蛋白参与膜泡的锚定
Rab蛋白在膜泡运输中的作用
NSF:N-乙基马来酰亚胺敏感因子,一种类似分子伴侣的ATP 酶α-SNAP:可溶性NSF 结合蛋白
2. SNARE配对是介导转运膜泡与靶膜融合的主要机制
图6-13 在供体膜和靶膜之间膜泡的锚定与融合模式图解
供体膜和靶膜之间膜泡的锚定与融合模式
信号假说与蛋白质分选信号
蛋白质分选的基本途径与类型
蛋白质向线粒体和叶绿体的分选
真核细胞中绝大多数蛋白质都是由核基因编码,在游离核糖体上起始合成
蛋白质的靶向转运(prtein targeting)或蛋白质分选(prtein srting)
真核细胞中,除少量蛋白质在线粒体和叶绿体内合成外,绝大多数蛋白质都是由核基因编码,或在游离的核糖体上合成,或在糙面内质网膜结合的核糖体上合成,然后转运至细胞特定部位,组装成结构与功能复合体,参与细胞生命活动的过程
一、信号假说与蛋白质分选信号
Fig. 2. Examples f directed transprt mediated by tpgenic signals. The figure shws a schematic cell with sme f its cmpartments, the rganelles. (A chlrplast is an rganelle that is present in plant cells but nt in animal cells). The rganelles have special functins and they are surrunded by membranes. Newly synthesized prteins are prvided with special “address tags”, signal sequences r tpgenic signals, which direct the prteins t a crrect place within the cell and allw them t crss the membranes f the rganelles. The signal itself cnsists f a chain f amin acids. It is an integral part f the prtein, and it is ften lcated at ne end f the prtein.
Fig. 1. “The signal hypthesis”. Prteins which are t be exprted ut f the cell are synthesized by ribsmes, assciated with the endplasmic reticulum. The genetic infrmatin frm DNA is transferred via messenger RNA (mRNA). This infrmatin determines hw the amin acids build up the prteins. First, a signal peptide is frmed as a part f the prtein. With the help f binding prteins, the signal peptide directs the ribsme t a channel in the endplasmic reticulum. The grwing prtein chain penetrates the channel, the signal peptide is cleaved, and the cmpleted prtein is released int the lumen f the endplasmic reticulum. The prtein is subsequently transprted ut f the cell.
The Nbel Prize in Physilgy r Medicine 1999
"fr the discvery that prteins have intrinsic signals that gvern their transprt and lcalizatin in the cell."
指导分泌性蛋白在粗面内质网上合成的决定因素包括:信号肽(signal peptide);信号识别颗粒(signal recgnitin particle,SRP);信号识别颗粒的受体(又称停泊蛋白,dcking prtein,DP);移位子(translcn, 或translcatr)
(1)信号肽(signal peptide)
位于蛋白质的N 端,一般由16~26 个残基组成;似乎没有严格的专一性包括疏水核心区、信号肽的C-端和N-端等3 部分原核细胞某些分泌性蛋白的N 端也具有信号序列
图6-1 信号肽的一级结构序列以血清白蛋白和HIV-1 型病毒的糖蛋白gp160 信号肽为例,显示出两者信号肽一级序列分别由疏水核心、C- 端和N- 端三个区域构成
(2)信号识别颗粒(SRP)
由6 种不同的蛋白质和一个由300 个核苷酸组成的 7S RNA 结合组成的一种核糖核蛋白复合体通常存在于细胞质基质中;既可与新生肽信号序列和核糖体大亚基结合,又可与内质网膜上SRP 受体结合
图6-2 信号识别颗粒(SRP)的结构示意图含有两个结构域,P54 蛋白是一种包含成簇甲硫氨酸残基的GTP酶,甲硫氨酸侧链与信号肽的疏水核心结合;当SRP 与信号肽结合后,SRP 的P9 和P14 蛋白复合体阻断新生肽链的翻译
(3)SRP 受体(Signal Recgnitin Particle Receptr)
内质网膜上的整合蛋白,由α 和β 亚基组成,可特异地与SRP 结合SRP 的p54 亚基和SRP 受体的α 亚基与GTP 结合时,会增进SRP/ 新生肽/ 核糖体复合物与SRP 受体结合的强度。
Dynamic behavir f SRP upn SR interactin
DOI: 10.1126/science.1124864
体外非细胞系统(cell free system)进行蛋白质合成实验,证实分泌性蛋白向rER(micrsme,微粒体)腔内的转运是同蛋白质翻译过程偶联进行的,这种分泌蛋白在信号肽引导下边翻译边跨膜转运的过程称为共翻译转运 (ctranslatinal translcatin)
蛋白质翻译过程与SRP、DP和微粒体的关系
在非细胞系统中蛋白质的翻译过程与SRP、DP 和微粒体的关系
* “+”和“-”分别代表反应混合物中存在(+)或不存在(-)该物质
(4)分泌性蛋白的合成与共翻译转运过程
图6-3 分泌性蛋白的合成与共翻译转运过程图解
分泌性蛋白的合成与其共翻译转运
Sec61 Prtein Translcatr
(5)内质网膜整合蛋白的信号序列
起始转移序列(start transfer sequence)内在停止转移锚定序列(internal stp-transfer anchr sequence, STA)内在信号锚定序列(internal signal anchr sequence, SA)
内质网膜整合蛋白的拓扑学类型
图6-4 内质网膜整合蛋白的拓扑学类型
单次跨膜蛋白插入转移机制:新生跨膜蛋白肽链中既有N端信号肽,又有停止转移信号(stp transfer signal)
整合膜蛋白的合成——单次跨膜
单次跨膜蛋白的插入转移机制:新生跨膜蛋白的肽链中没有N 端信号肽,只有内部信号序列(internal signal sequence)。内部信号序列的转位方向主要取决于其侧翼氨基酸残基电荷的分布。一般而言,带正电荷的氨基酸残基一侧朝向细胞质基质一侧
多次跨膜蛋白的插入转移机制:多次跨膜蛋白肽链中,常常有两个或两个以上的疏水性开始转移信号序列和停止转移信号序列。一般认为,多次跨膜蛋白是以内部信号序列(internal signal sequence)作为其开始转移信号
整合膜蛋白的合成——多次跨膜
蛋白质自身所携带的分选信号需要被互补的分选受体所识别,然后在分选受体帮助下,引导蛋白质转运到目的地
2. 蛋白质分选信号序列
导肽(leader peptide):指导在细胞质基质中合成的蛋白质进入线粒体、叶绿体及过氧化物酶体的信号序列
表6-2 指导蛋白质从细胞质基质转运到细胞器的靶向序列的主要特征
2. 蛋白质分选信号序列(signal sequence)
两类分选信号一类为连续的氨基酸序列,一般长15~60个氨基酸残基。完成分选过程后,有些这类信号序列会被特异性酶从成熟蛋白质上切除一类为三维结构的信号斑,在蛋白折叠完成后,其表面原子按一定的间结构排布形成。形成信号斑的氨基酸残基可能在线性氨基酸序列上相距很远。信号斑会保留在成熟蛋白质中不被切除
途径(1)翻译后转运途径(2)共翻译转运途径类型(1)蛋白质的跨膜转运(transmembrane transprt)(2)膜泡运输(vesicular transprt)(3)选择性的门控转运(gated transprt)(4)细胞质基质中蛋白质的转运
二、蛋白质分选转运的基本途径与类型
真核细胞蛋白质分选的主要途径与类型
图6-5 真核细胞蛋白质分选的主要途径与类型
三、蛋白质向线粒体和叶绿体的分选
转运到线粒体、叶绿体和过氧化物酶体等中的蛋白需要多个不同靶向序列(targeting sequences);细胞器蛋白最终是定位在不同膜上还是不同基质空间,除N- 端不同转运肽外,还需要其他空间定位信号序列通过翻译后转运途径进入线粒体、叶绿体和过氧化物酶体等细胞器的蛋白,必须在分子伴侣帮助下解折叠或维持非折叠状态,蛋白质输入这些细胞器通常是需要能量的过程
1. 蛋白质从细胞质基质输入到线粒体
蛋白质从细胞质基质输入到线粒体基质蛋白质以3 种途径从细胞质基质输入到线粒体内膜线粒体蛋白通过2 种途径从细胞质基质输入到线粒体膜间空间
图6-6 通过后翻译转运途径,核基因编码的线粒体蛋白的转运A. 蛋白质从细胞质基质输入到线粒体基质(图左上);B. 线粒体蛋白通过3 种途径从细胞质基质输入到线粒体内膜(图右);C. 线粒体蛋白通过2 种途径从细胞质基质输入到线粒体膜间空间(图左下)
外膜蛋白内膜蛋白叶绿体基质蛋白类囊体膜蛋白类囊体腔蛋白
2. 蛋白质从细胞质基质向叶绿体的分选
图6-7 通过后翻译转运途径,叶绿体蛋白从细胞质基质输入到类囊体腔注:Tm/Tim:Translcn f the uter/inner membrane
叶绿体蛋白从细胞质基质输入到类囊体腔
COP Ⅱ 包被膜泡的装配及运输
COP I 包被膜泡的装配与运输
网格蛋白/接头蛋白包被膜泡的装配与运输
转运膜泡与靶膜的锚定和融合
The Nbel Prize in Physilgy r Medicine 2013
"fr their discveries f machinery regulating vesicle traffic, a majr transprt system in ur cells."
James E. Rthman
James E. Rthman discvered that a prtein cmplex (pictured in range) enables vesicles t fuse with their target membranes. Prteins n the vesicle bind t specific cmplementary prteins n the target membrane, ensuring that the vesicle fuses at the right lcatin and that carg mlecules are delivered t the crrect destinatin.
Randy W. Schekman
Randy W. Schekman discvered genes encding prteins that are key regulatrs f vesicle traffic. Cmparing nrmal with genetically mutated yeast cells in which vesicle traffic was disturbed, he identified genes that cntrl transprt t different cmpartments and t the cell surface.
secretin-defective mutant strain (sec).
Thmas C. Südhf
Thmas C. Südhf studied hw signals are transmitted frm ne nerve cell t anther in the brain, and hw calcium cntrls this prcess. He identified mlecular machinery (pictured in purple) that senses calcium ins (Ca2+) and triggers vesicle fusin, thereby explaining hw tempral precisin is achieved and hw signaling substances can be released frm the vesicles n cmmand.
图6-8 蛋白质的分泌与胞吞途径概观1. 在rER 合成的蛋白质, 通过共翻译转运途径跨膜运输;2. 内质网出芽,形成转运泡并与高尔基体融合,形成cis- 高尔基网状结构;3. 从cis- 高尔基膜囊和cis- 高尔基体顺面网状结构到rER 的逆向运输;4. 高尔基膜囊从cis 面→ trans 面成熟递进(非膜泡过程);5. 从高尔基体后期膜囊→早期膜囊的逆向运输;6. 组成型分泌;7. 调节型分泌;8. 分选到溶酶体;9. 胞吞途径
Fig. A “rad-map” f the secretry and endcytic pathways
三种不同类型的包被膜泡
图6-9 在细胞合成- 分泌与内吞途径中三种不同的主要膜泡运输方式COP II(cat prtein II)包被泡、COPI(cat prtein I)包被泡和网格蛋白/ 接头蛋白(clathrin/adaptr prtein)包被泡,它们分别介导不同的膜泡转运途径
COPⅡ包被膜泡介导从内质网到高尔基体的顺向运输。COPⅠ包被膜泡介导细胞内膜泡逆向运输,负责从高尔基体反面膜囊到高尔基体顺面膜囊,以及从高尔基体CGN区到内质网的膜泡转运。网格蛋白/ 接头蛋白包被膜泡介导几种蛋白质分选途径,包括从高尔基体TGN向胞内体、溶酶体、黑(色)素体、血小板囊泡或植物细胞液泡的运输。另外,在受体介导的胞吞途径中还负责将物质从细胞表面运往胞内体转而到溶酶体的运输
蛋白质转运中涉及的3 种包被膜泡的特征比较
表8-3 蛋白质转运中涉及的三种包被膜泡的特征比较
COP Ⅱ 包被膜泡介导细胞内顺向运输(antergrade transprt),即负责从内质网到高尔基体的物质运输COP Ⅱ 包被蛋白组分小分子GTP 结合蛋白Sar1Sec23/Sec24 复合物Sec13/Sec31 复合物以及大的纤维蛋白Sec16
二、COP II包被膜泡的装配及运输
COP Ⅱ 包被膜泡的装配与去装配
图6-10 Sar1 蛋白在Cp Ⅱ 包被膜泡装配与去装配中作用模型1. Sar1 与膜结合,GTP 交换;2. COP Ⅱ包被装配;3. GTP 水解;4. COP Ⅱ包被去装配
Sar1 蛋白在CpⅡ包被膜泡装配与去装配中作用
细胞质中可溶性 Sar1- GDP 与ER 膜蛋白Sec12(鸟苷酸交换因子)相互作用, 催化GTP 置换GDP 形成Sar1-GTP,GTP 的结合引发 Sar1 构象改变暴露出疏水N 端并插入ER 膜,膜结合Sar1 对包被蛋白进一步装配起募集者作用
COP Ⅱ 包被膜泡装配
小分子GTP 结合蛋白Sar1
Sec23 是GTP酶活化蛋白(GTPase-activating prtein, GAP),能够提高Sar1水解GTP 的酶活性。特别是当Sec31 被招募到复合体中后,能够提高Sec23 作为Sar1的GAP 活性达10 倍,从而使成熟的COPII包被膜泡很容易发生去装配
COPⅡ包被膜泡的去装配
图6-11 不同类型的膜泡运输A. 内质网与高尔基体之间,分别由COPⅡ和COPⅠ膜泡介导蛋白质顺向和逆向转运:步骤1~3 表示由COPⅡ包被膜泡介导的顺向运输,内质网膜出芽,可溶性包被蛋白复合物聚合形成膜泡包被;v-SNAREs 和其它被转运的膜蛋白和可溶性蛋白(与芽泡膜上受体结合),通过与包被蛋白的相互作用被包装在膜泡内,包被复合物解聚再循环利用,v-SNAREs 蛋白在脱包被膜泡表面得以暴露,以利于同靶膜的融合。步骤4~6 表示由COPⅠ包被膜泡介导的反向运输,膜脂双分子层、某些蛋白如v-SNAREs 和错误分选的内质网驻留蛋白,从cis- 高尔基网状区转运到内质网。B. KDEL 受体在从高尔基体回收内质网腔驻留蛋白中的作用:步骤1~2 表示内质网腔中高水平存在的蛋白C- 端具有KDEL 信号,被包装到COPⅡ包被膜泡,并转运到高尔基体;步骤3~4 表示内质网逃逸蛋白通过COPⅠ膜泡介导的反向运输得以回收,注意,COPⅡ、COPⅠ和cis- 高尔基网状区膜上均有识别与结合KDEL 信号的受体,信号与受体的亲和力受到pH 高低的影响,低pH 值促进结合,高pH 值有利于释放。C. 在供体膜内质网出芽及其转运蛋白的包装示意图:通过募集小分子GTP 结合蛋白,引发供体膜出芽,然后,胞质中的包被蛋白复合物与被转运的膜蛋白胞质结构域结合,其中有些膜蛋白也作为腔内可溶性蛋白的受体
三、COP I 包被膜泡的装配及运输
COP I 包被膜泡介导细胞内膜泡逆向运输(retrgrade transprt),负责从高尔基体反面膜囊到高尔基体顺面膜囊以及从高尔基体顺面网状区到内质网的膜泡转运含有7 种不同的蛋白质亚基和一种调节膜泡转运的 GTP 结合蛋白ARF(ADP Ribsylatin Factr 1 )ARF 也是一种结合 GDP/GTP 转换的分子开关调控蛋白
Cat assembly drives vesicle frmatin
KDEL 受体主要定位在高尔基体TGN 区、COPⅡ和COPⅠ包被膜泡的膜上,它们能识别并结合KDEL 分选信号。如果在内质网发生错误包装和转运,由于COPⅠ膜泡上也有KDEL 受体,所以也能保证逃逸蛋白被内质网回收
COP Ⅱ和COP Ⅰ包被膜泡介导内质网与高尔基体之间蛋白质顺向和逆向转运
①~③表示由COPⅡ包被膜泡介导的顺向运输,内质网膜出芽,可溶性包被蛋白复合物聚合形成膜泡包被;v-SNARE,以及其他被转运的膜蛋白和可溶性蛋白(与芽泡膜上受体结合)通过与包被蛋白的相互作用被包装在膜泡内,包被复合物解聚再循环利用,v-SNARE蛋白在脱包被膜泡表面得以暴露,以利于同靶膜的融合。④~⑥ 表示由COPⅠ包被膜泡介导的逆向运输,膜脂双分子层、某些蛋白质(如v-SNARE和错误分选的内质网驻留蛋白)从高尔基体CGN区转运回内质网
网格蛋白/ 接头蛋白包被膜泡介导的蛋白质分选途径从高尔基体TGN向胞内体或向溶酶体、 黑(色)素体、血小板囊泡和液泡的运输受体介导的胞吞途径中负责将物质从细胞表面运往胞内体转而到溶酶体的运输
四、网格蛋白/接头蛋白包被膜泡的装配与运输
图6-12 网格蛋白、多角形网格包被结构及其的发动蛋白介导的网格蛋白/AP 包被小泡的形成示意图A. 三腿网格蛋白分子,3 条重链,3 条轻链;B. 在体外缺乏膜泡的情况下自组装的网格蛋白网格包被(基于PDB 数据库1XI4 结构绘制)。C. 发动蛋白介导的网格蛋白/AP 包被小泡的形成示意图
网格蛋白及其包被膜泡的形成
胞吞过程中网格蛋白/接头蛋白包被膜泡的装配
ARF 也参与网格蛋白/ 接头蛋白包被的起始装配(细胞膜—胞内体 不需要)发动蛋白(dynamin)具有GTPase 活性,导致网格蛋白/ 接头蛋白包被膜泡从供体膜断裂并释放
发动蛋白在网格蛋白包被膜泡形成中的作用
五、转运泡与靶膜的锚定与融合
① 供体膜的出芽、装配和断裂,形成不同的包被转运膜泡② 在细胞内由马达蛋白驱动、以微管为轨道的膜泡运输③ 转运膜泡与特定靶膜的锚定和融合
可溶性N-乙基马来酰亚胺敏感因子结合蛋白受体SNARE (sluble N ethyl maleimide-sensitive factr attachment prtein receptr)
Rab (ras-related in brain)
Ras (rat sarcma, Ras)
v-SNARE (vesicle SNARE)
t-SNARE (target SNARE)
1. Rab蛋白参与转运膜泡与靶膜的识别与锚定
胞质中Rab 蛋白在特异性鸟苷酸交换因子(GEF)催化下,Rab-GDP 转换为Rab-GTP,构象改变致使其通过类异戊二烯基团插入转运膜泡表面,与靶膜上Rab 效应器结合蛋白相互作用,从而使转运膜泡被锚定在靶膜上
Rab 蛋白参与膜泡的锚定
Rab蛋白在膜泡运输中的作用
NSF:N-乙基马来酰亚胺敏感因子,一种类似分子伴侣的ATP 酶α-SNAP:可溶性NSF 结合蛋白
2. SNARE配对是介导转运膜泡与靶膜融合的主要机制
图6-13 在供体膜和靶膜之间膜泡的锚定与融合模式图解
供体膜和靶膜之间膜泡的锚定与融合模式
相关课件
高考生物一轮复习第7讲物质跨膜运输的实例物质跨膜运输的方式课件: 这是一份高考生物一轮复习第7讲物质跨膜运输的实例物质跨膜运输的方式课件,共46页。PPT课件主要包含了-3-,必备知识梳理,关键能力突破,-4-,-5-,-6-,-7-,-8-,-9-,-10-等内容,欢迎下载使用。
人教版 (2019)二 酶的特性背景图ppt课件: 这是一份人教版 (2019)二 酶的特性背景图ppt课件,共20页。
第04章 物质的跨膜运输课件PPT: 这是一份第04章 物质的跨膜运输课件PPT,共60页。PPT课件主要包含了本章主要内容,胞吞作用与胞吐作用,第一节,电压门控通道,第二节,第三节,本章内容提要等内容,欢迎下载使用。











