



还剩52页未读,
继续阅读
第09章 细胞核与染色质课件PPT
展开
这是一份第09章 细胞核与染色质课件PPT,共60页。PPT课件主要包含了本章主要内容,核被膜,染色质,染色质的复制与表达,染色体,核仁与核体,核基质,第一节,核膜结构,第二节等内容,欢迎下载使用。
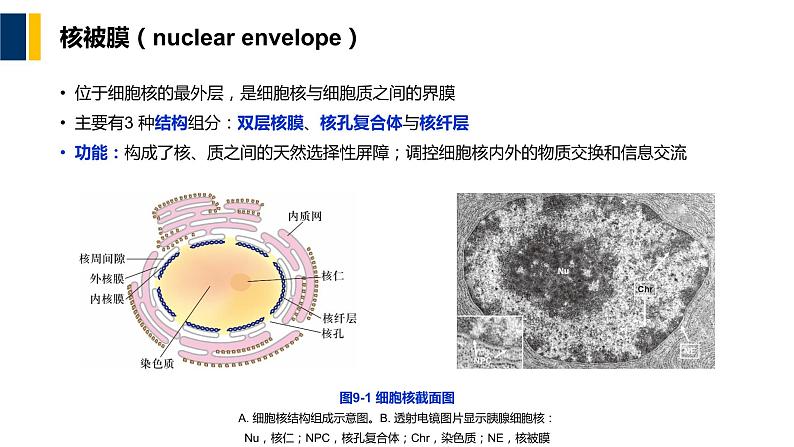
核被膜(nuclear envelpe)
位于细胞核的最外层,是细胞核与细胞质之间的界膜主要有3 种结构组分:双层核膜、核孔复合体与核纤层功能:构成了核、质之间的天然选择性屏障;调控细胞核内外的物质交换和信息交流
图9-1 细胞核截面图A. 细胞核结构组成示意图。B. 透射电镜图片显示胰腺细胞核:Nu,核仁;NPC,核孔复合体;Chr,染色质;NE,核被膜
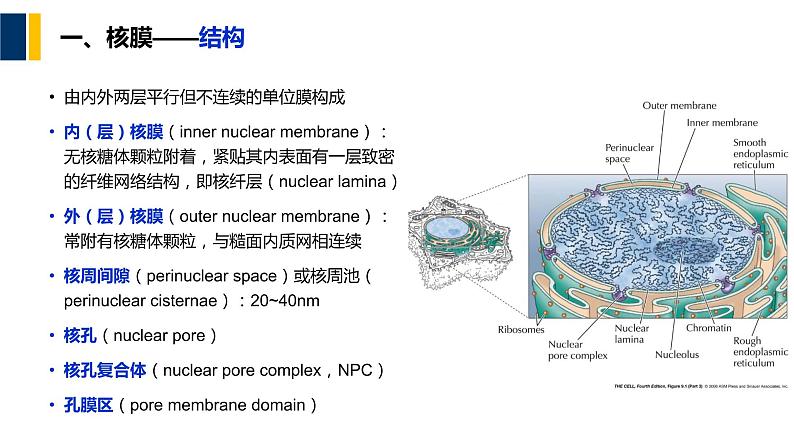
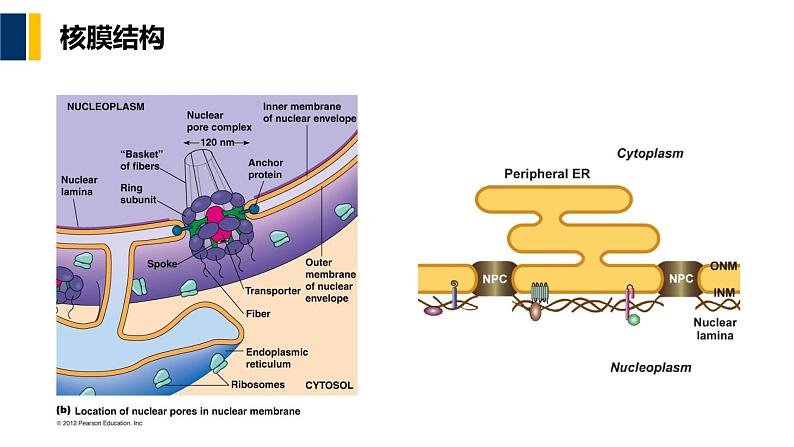
由内外两层平行但不连续的单位膜构成内(层)核膜(inner nuclear membrane):无核糖体颗粒附着,紧贴其内表面有一层致密的纤维网络结构,即核纤层(nuclear lamina)外(层)核膜(uter nuclear membrane):常附有核糖体颗粒,与糙面内质网相连续核周间隙(perinuclear space)或核周池(perinuclear cisternae):20~40nm核孔(nuclear pre)核孔复合体(nuclear pre cmplex,NPC)孔膜区(pre membrane dmain)
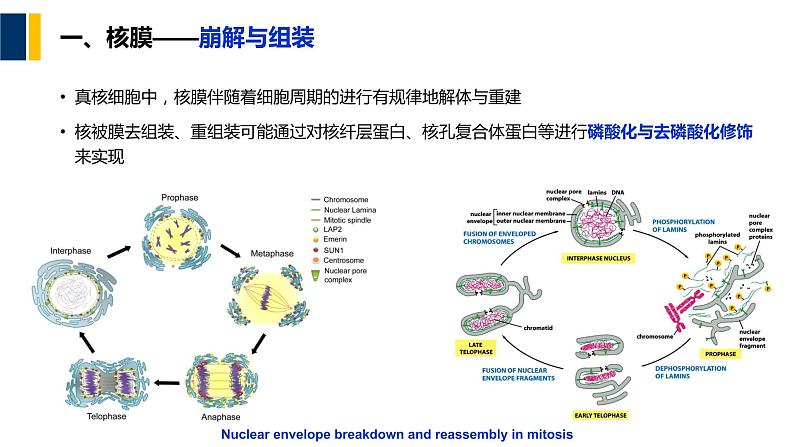
一、核膜——崩解与组装
真核细胞中,核膜伴随着细胞周期的进行有规律地解体与重建核被膜去组装、重组装可能通过对核纤层蛋白、核孔复合体蛋白等进行磷酸化与去磷酸化修饰来实现
Nuclear envelpe breakdwn and reassembly in mitsis
核孔复合体是一个相对独立的复杂结构;间期细胞核普遍存在核孔复合体在核被膜上的数量、分布密度与分布形式随细胞类型、细胞核的功能状态的不同而有很大的差异研究核孔复合体形态结构的经典方法主要有三种:树脂包埋超薄切片技术、负染色技术、冷冻蚀刻技术
图9-2 用冷冻蚀刻电镜技术(A)和高分辨率扫描电镜(B)显示的核孔复合体(NPC)
胞质环(cytplasmic ring)核质环(nuclear ring)辐(spke):柱状亚单位;腔内亚单位;环带亚单位中央栓(central plug),中央颗粒 (central granule)
图9-3 核孔复合体结构模型
(一)结构模型—— “fish-trap”
Nature Reviews Mlecular Cell Bilgy 11, 490-501 (July 2010) | di:10.1038/nrm2928
(二)组成成分——核孔蛋白 (nucleprin,Nup)
gp210:结构性跨膜蛋白介导NPC与核被膜连接在内、外核膜融合形成核孔中起重要作用在核质交换中起作用p62:功能性Nup疏水性N 端区: 具有FXFG重复序列C 端区:具有疏水性的7肽重复序列
双功能、双向性的亲水性核质交换通道双功能表现在它有两种运输方式:被动扩散与主动运输双向性表现在既介导蛋白质的入核转运,又介导RNA、核糖核蛋白颗粒(RNP)的出核转运
1.通过核孔复合体的被动扩散
图9-4 通过核孔复合体物质运输的功能示意图
2.核孔复合体的主动运输
具有高度选择性对运输颗粒大小的限制是一个信号识别与载体介导的过程,需要消耗ATP 能量,并表现出饱和动力学特征双向性:核输入与核输出
亲核蛋白(karyphilic prtein)入核转运机制
亲核蛋白:在细胞质内合成后,需要或能够进入细胞核内发挥功能的一类蛋白质包括:在一个细胞周期中一次性地被转运到核内,并一直停留在核内行使功能的亲核蛋白,如组蛋白、核纤层蛋白等;需要穿梭于核质之间进行功能活动的亲核蛋白,如imprtin核定位序列(nuclear lcalizatin sequence)或核定位信号(nuclear lcalizatin signal,NLS):引导亲核蛋白入核,富含碱性氨基酸残基,如Lys、Arg,还常含有Pr;可以是一段连续的序列,也有分成两段存在;受体为imprtin;完成核输入后不被切除核输出信号(nuclear exprt signal,NES):引导RNP输出细胞核,受体为exprtinRan蛋白:属G蛋白,调节货物复合体的解体或形成
注射核质蛋白及核质蛋白组分的细胞定位
A classic experiment that used several frms f radiactive nucleplasmin(核质蛋白), which is a large pentameric prtein invlved in chrmatin assembly. Either the intact prtein r the nucleplasmin heads, tails, r heads with a single tail were injected int the cytplasm f a frg cyte r int the nucleus. All frms f nucleplasmin, except heads, accumulated in the nucleus when injected int the cytplasm, and all frms were retained in the nucleus when injected there.A. 核质蛋白哪部分负责其在核内定位?B.该实验如何证明其入核是由核定位信号指导的主动运输,而不是被动扩散?
核定位信号与Ran蛋白
亲核蛋白通过核孔复合体的转运:结合(binding)与转移(translcatin)
亲核蛋白NLS 识别imprtin α,与可溶性受体imprtin α/β 异二聚体结合,形成转运复合物在imprtin β 介导下,转运复合物与核孔复合体的胞质纤维结合转运复合物通过改变构象的核孔复合体从胞质面被转移到核质面转运复合物在核质面与Ran-GTP 结合,并导致复合物解离,亲核蛋白释放受体亚基与结合的Ran 返回胞质,在胞质内Ran-GTP 水解形成Ran-GDP 并与imprtin β 解离,Ran-GDP返回核内再转换成Ran-GTP 状态
图9-5 亲核蛋白从细胞质向细胞核输入的过程示意图
NPC介导双向核质转运
三、核纤层 nuclear lamina
3 种核纤层蛋白:lamin A、B 、C核纤层蛋白本身形成纤维状网络结构功能:结构支撑;调节基因表达;调节DNA 修复;与细胞周期有关
图9-6 冷冻蚀刻电镜技术显示核纤层的纤维结构
In HGPS patients the cell nucleus has dramatically aberrant mrphlgy (bttm, right) rather than the unifrm shape typically fund in healthy individuals (tp, right)
Hutchinsn-Gilfrd早衰综合征(HGPS):核纤肽A的C端缺失50个氨基酸
染色质 (chrmatin)
间期细胞核内由DNA、组蛋白、非组蛋白及少量RNA 组成的线性复合结构,是间期细胞遗传物质存在的形式细胞内基因存在与发挥功能的结构基础与基因组直接相关的细胞活动都是在染色质水平进行,如DNA 复制、基因转录、同源重组、DNA 修复,包括转录偶联的修复(transcriptin cupled repair)以及DNA 和组蛋白的各种修饰
图9-7 DNA 复制、转录和重组都是在染色质水平进行
一、染色质DNA——基因组大小比较
基因组(genme):某一生物的细胞中储存于单倍染色体组中的总遗传信息真核生物基因组DNA 的含量比原核生物高得多最小最简单的细胞支原体有迄今发现的能独立生活的有机体的最小基因组
Figure 1-37 Mlecular Bilgy f the Cell, Fifth Editin (© Garland Science 2008)
一、染色质DNA——基因组DNA 类型
蛋白编码序列编码rRNA、tRNA、snRNA和组蛋白的串联重复序列含有重复序列的DNA未分类的间隔DNA
DNA二级结构:两条多核苷酸链反向平行盘绕所生成的双螺旋结构分3 种:B 型DNA(右手双螺旋),“经典”的Watsn-Crick 结构;A 型DNA(B 型DNA 变构形式,右手双螺旋);Z 型DNA (B 型DNA 变构形式,左手螺旋)大沟的特征在遗传信息表达过程中起关键作用
组蛋白(histne)与DNA 结合没有序列特异性非组蛋白(nnhistne)与特定 DNA 序列或组蛋白相结合
属碱性蛋白质,富含带正电荷的Arg 和Lys 等碱性氨基酸,可以和酸性DNA 紧密结合,一般不要求特殊核苷酸序列核小体组蛋白 H2A、H2B、H3 和H4没有种属及组织特异性,进化上十分保守H1 组蛋白有一定的种属和组织特异性
(一)组蛋白(histne)
(二)非组蛋白(nnhistne)
序列特异性 DNA结合蛋白 特性多样性识别DNA具有特异性具有功能多样性
凝胶延滞实验(gel retardatin assay)
三、核小体(nuclesme)
核小体:染色质组装的基本结构单位染色质结构的“串珠”模型
图9-8 透射电镜显示串珠状11 nm 的核小体结构
透射电镜显示串珠状11 nm 的核小体结构
Figure 4-22 Mlecular Bilgy f the Cell (© Garland Science 2008)
盐溶液处理染色质11nm
非特异性微球菌核酸酶(micrcccal nuclease)消化染色质,DNA 被降解成约200 bp 的片段
X 射线晶体衍射揭示核小体三维结构
图9-9 由X 射线晶体衍射所揭示的人类细胞核小体三维结构A. 通过DNA 超螺旋中心轴所显示的核小体核心颗粒8 个组蛋白分子的位置。B. 垂直于中心轴的角度所见到的核小体核心颗粒的盘状结构
SV40 微小染色体(minichrmsme)分析
Virlgy, 2009, 384(2): 352-359
包括200 bp 左右的DNA 超螺旋和一个组蛋白八聚体以及一个分子的组蛋白H1组蛋白八聚体构成核小体的盘状核心颗粒,由4 个异二聚体组成,包括两 个H2A • H2B 和两个H3 • H4
146/147 bp:盘绕组蛋白八聚体1.75 圈166/167 bp:组蛋白H1 结合额外20 bp DNA,锁住核小体DNA的进出端,稳定核小体200 bpLinker DNA:相邻核小体之间;典型长度为60 bp,不同物种变化值为0~80 bp 不等
图 9-10 核小体的结构要点示意图
组蛋白与DNA 之间的相互作用主要是结构性的,基本不依赖于核苷酸的特异序列核小体沿DNA 的定位受不同因素的影响非组蛋白与DNA 特异性位点的结合,可影响邻近核小体的相位(psitining)富含AT 的DNA片段优先存在于DNA 双螺旋的小沟,而富含GC 的DNA 片段优先存在于DNA 双螺旋的大沟
Figure 4-23 Mlecular Bilgy f the Cell (© Garland Science 2008)
用稀酸或高盐溶液处理染色质,可使组蛋白与 DNA 解离,如何解释?
组蛋白与 DNA 之间的结合依靠的是组蛋白带正电的碱 性基团与 DNA 带负电荷的磷酸基团之间的静电引 力,如果用稀酸处理复合物,则磷酸基团质子化而 失去所带的负电荷,复合物解离。如果用高盐溶液 处理复合物,则阳离子与磷酸基团结合而取代了组 蛋白,导致组蛋白与 DNA 解离。
人体细胞46条染色体含DNA约6×109 bp,总长2米,如何计算出来的?
裸露DNA 组装直径30 nm 螺线管H3 • H4 两个异二聚体结合两个H2A • H2B 异二聚体加入组蛋白去乙酰化,H1组蛋白结合伴随着核小体折叠6 个核小体组成一个螺旋旋或由其他组装方式形成一个螺线管结构进一步折叠事件使染色质在核中形成确定的结构
图9-11 DNA 组装成染色质的过程及各阶段的协助组装因子
(一)染色质组装的前期过程
一级结构:核小体二级结构:螺线管三级结构:超螺线管四级结构:染色单体
图9-12 30 nm 染色质纤维的三维结构
(二)染色质组装的多级螺旋模型
(三)染色质组装的放射环结构模型
电镜:非组蛋白构成染色体骨架(chrmsmal scaffld)和与骨架相连的无数DNA 侧环染色体的骨架-放射环结构模型(scaffld radial lp structure mdel):30 nm 的染色线折叠成环,沿染色体纵轴,由中央向四周伸出,构成放射环
染色质具有高度复杂的结构
图9-13 基因组结构和染色质组装一览图
五、染色质类型 (一)常染色质与异染色质
常染色质(euchrmatin):间期细胞核内染色质纤维折叠压缩程度低,相对处于伸展状态,用碱性染料染色时着色浅异染色质 (heterchrmatin):间期细胞核中染色质纤维折叠压缩程度高,处于聚缩状态,用碱性染料染色时着色深结构异染色质或组成型异染色质(cnstitutive heterchrmatin):整个细胞周期均处于聚缩状态,没有较大变化;在中期染色体上多定位于着丝粒区、端粒、次缢痕等;由相对简单、高度重复的DNA 序列构成;具有显著的遗传惰性;为晚复制、早聚缩兼性异染色质 (facultative heterchrmatin):在某些细胞类型或一定的发育阶段,原来的常染色质聚缩,并丧失基因转录活性,变为异染色质;如巴氏小体(Barr bdy)
Barr Bdies: heterchrmatinized X-chrmsmes
异染色质或常染色质之间随着发育时期或细胞周期的变化可能相互转化异染色质与常染色质之间的转变常常需要伴随着一些组蛋白与DNA 修饰
图9-14 常染色质与异染色质转变过程中伴随的组蛋白修饰的变化
(二)常染色质与异染色质间的转变
活性染色质(active chrmatin)具有转录活性具有疏松的染色质结构非活性染色质(inactive chrmatin) 没有转录活性
(三)活性染色质与非活性染色质
1.活性染色质对DNase Ⅰ超敏感
活化染色质对DNaseⅠ的优先敏感性(preferential sensitivity)是可转录染色质一个基本特征活化染色质对DNaseⅠ的优先敏感性与HMG14 和HMG17两种非组蛋白有关,但不是唯一原因DNaseⅠ超敏感位点(DNaseⅠ hypersensitive site)的存在是活性染色质的特点
2.活性染色质的蛋白组成与修饰变化
活性染色质很少有组蛋白H1 与其结合;组蛋白乙酰化程度高,且H2B很少被磷酸化H2A 在活性染色质中很少有变异形式存在;H3 变种H3.3 只在活跃转录的染色质中出现HMG14 和HMG17 只存在于活性染色质中,与DNA 结合活性染色质标志:H3 N 端第4 个赖氨酸甲基化,第9 和14 个赖氨酸乙酰化以及第10 个丝氨酸磷酸化;非活性染色质标志:H3 N 端第9 个赖氨酸甲基化而不是乙酰化
图9-15 H3 组蛋白修饰与染色质活性的关系
染色质的三维动态分布与细胞ID
一、染色质的复制与修复
%20Glasgw%20June%202010/Image1.gif
S 期染色质完全复制不仅需要基因组DNA 的复制,也需要把复制好的DNA组装成染色质新复制的DNA 主要通过两种途径组装成染色质:第一,在复制叉的移动期间,父代的核小体核心颗粒与DNA 分离,到该段DNA 复制完成,父代的核小体核心颗粒直接转移到两条子链DNA 的一条上第二,染色质组装因子利用刚刚合成的、乙酰化的组蛋白介导核小体在复制DNA 上组装
图9-16 异染色质复制过程示意图
高度浓缩的染色质结构对基因组DNA(染色质DNA)具有重要的保护作用染色质DNA 随着细胞周期的变化会暴露或部分暴露于其周围环境中。基因组DNA 会受到细胞内外化学的与物理的因素的作用产生突变细胞中存在一整套的DNA 修复机器来应对各种各样的DNA 突变染色质修复(chrmatin restratin):修复后的DNA 组装成染色质的过程细胞一旦丧失了DNA 修复的部分或全部能力,基因组就会变得不稳定。基因组的不稳定性是直接导致肿瘤发生的重要原因之一
(二)染色质的修复与基因组稳定性
Fig. Chrmatin mdificatins and their cnsequences in respnse t lesins that are repaired by NER
二、染色质的激活与失活
染色质结构改变与基因活化的关系的研究主要集中在3 个方面:如何形成活性染色质,以便RNA 聚合酶能起始转录具有转录活性的染色质结构域如何与周围的非活性区域隔离RNA 聚合酶如何通过与染色质模板结合进行转录
染色质是一个动态、可塑的蛋白质与核酸组成的复合体调控蛋白结合到染色质DNA 的特定的位点上时,不管是在核小体间还是在一个核小体内,染色质都很容易被引发二级结构的改变。这些改变使得其他的一些结合位点与调控蛋白的结合变得要么更加容易,要么更加困难
图9-17 核小体变构因子通过改变核小体的相位而调节染色质的活性
(一)染色质的激活——DNA 结构与核小体相位的变化
核心组蛋白的赖氨酸残基乙酰化组蛋白H1 的磷酸化不同组蛋白修饰之间的关系
(一)染色质的激活——组蛋白的修饰
核心组蛋白的赖氨酸残基乙酰化(acetylatin)
图9-18 乙酰化和去乙酰化对染色质活性的影响A. 乙酰化和去乙酰化的位置。B. 导致染色质的变化
图9-19 酵母组蛋白乙酰化与去乙酰化控制转录的作用机制URS: 上游抑制序列(upstream repressin sequence);UAS: 上游激活序列;RD: 抑制结构域;AD: 激活结构域。
高速泳动族非组蛋白HMG1 和 HMG2 有A、B 和C 3 个结构域HMG 结构域蛋白结合在DNA 双螺旋的小沟中与DNA 弯折和DNA-蛋白质复合体高级结构形成有关
HMG (high-mbility grup)
(一)染色质的激活——HMG 蛋白的影响
图9-20 哺乳动物X 染色体失活模型Xic:X 染色体失活中心;Xist RNA:X 染色体失活特异性转录物;RNA(X-inactivatin specific transcript RNA)
(二)染色质的失活——X 染色体失活
活性基因转位到异染色质区附近时会失活的现象称为位置花斑效应(psitin effect variegatin)
(二)染色质的失活——位置花斑效应
三、染色质与基因表达调控
Circulatin Research. 2010; 107: 324-326
(一)以染色质为模板的转录
染色质是否处于活化状态(呈疏松或紧密结构)是决定RNA 聚合酶能否有效行使转录功能的关键转录起始伴随着染色质上某一基因调节序列内部或者周围的结构改变RNA聚合酶被认为是用“核小体犁”(nuclesme plw)来解除组蛋白和DNA 间的相互作用
图9-21 酵母激活因子及辅激活子有顺序地相互作用于HO 基因在染色质水平调节其表达
通用转录因子与结合RNA 聚合酶的核心启动子位点结合特异转录因子与特异基因的各种调控位点结合,促进或阻遏基因的转录DNA 结合域结合特异的DNA 序列激活结构域 激活转录
(二)转录因子介导的基因表达调控
转录抑制因子结合于特定的调节DNA 序列,抑制结合区附近基因转录DNA 结合域抑制结构域
图9-22 基因表达的时空特异性图中红色显示的是在果蝇的翅膀成虫盘(wing disc)中,dpp-GAL4 介导的Uif 的表达在前后分界处。绿色是在同一个盘中的背腹分界处的Wg 的表达。dpp:decapentaplegic;Uif:uninflatable;wg:Wingless
(三)DNA 甲基化介导的基因表达调控
DNA 甲基化(DNA methylatin)与基因表达的阻遏有关DNA 甲基化对维持基因长期处于非活化状态起重要作用。非活跃转录基因的DNA 甲基化程度普遍高于活跃转录的基因甲基化作用通过两种方式抑制转录:一是干扰转录因子对DNA 结合位点的识别,二是将转录激活因子识别的DNA 序列转换为转录抑制因子的结合位点DNA 甲基化在基因组印记(genmic imprinting)中具有重要作用
Fig. Regulatin f DNA methylatin f gene expressin
(四)组蛋白修饰介导的基因表达调控
转录因子能够调节组蛋白核心的结构,改变核小体和染色质的紧密程度,影响通用转录因子和RNA 聚合酶对启动子的结合,调控基因表达转录激活因子通常有利于那些导致染色质或组蛋白结构松散的蛋白质复合物发挥作用,如组蛋白乙酰化酶转录抑制因子通常会加强那些促进染色质结构紧密的蛋白质的作用,如组蛋白去乙酰化酶
( 五)染色质与表观遗传
表观遗传的变化是通过组蛋白和DNA 的不同修饰而实现的表观遗传修饰可以遗传但富于变化,通过改变染色质的结构而改变基因组信息的表达可改变染色质结构的途径主要包括:组蛋白密码子(histne cde)、染色质重塑(chrmatin remdeling)、组蛋白异型体(histne variants)、DNA 甲基化和非编码RNA(nncding RNAs)的作用
四、染色质的三维动态分布与细胞ID
染色质在细胞核内的分布不是均匀的,也不是静止不变的,而是存在时间和空间上的特异性特异分布的特性与细胞内的基因表达相关,更与细胞的状态直接相关细胞的身份(identity)有可能根据某些标志性DNA 序列的三维分布建立根类细胞ID 标准化信息库
Fig. Schematic representatin f hierarchical chrmatin rganizatin in plants
染色体是细胞在有丝分裂(或减数分裂)时遗传物质存在的特定形式,是间期细胞染色质结构紧密组装的结果由两条相同的姐妹染色单体(chrmatid)构成,彼此以着丝粒 (centrmere)相连根据着丝粒在染色体上所处的位置,分为4 种类型
图9-23 根据着丝粒位置进行的染色体分类图示
动粒结构域(kinetchre dmain) 内板(inner plate)中间间隙 (middle space)外板 (uter plate)纤维冠(fibruscrna)中央结构域(central dmain)配对结构域(pairing dmain)
图9-24 着丝粒的结构域组织
(二)染色体的其他主要结构
次缢痕(secndary cnstrictin)核仁组织区(nuclelar rganizing regin,NOR)随体(satellite)端粒(telmere)
二、染色体的功能元件(functinal element)
构成染色体DNA 的3 种关键序列(key sequence)称为染色体DNA 的功能元件DNA 复制起点着丝粒端粒“人造微小染色体”(artificial minichrmsme)
图9-25 真核细胞染色体的3 种功能元件示意图
(一)自主复制DNA序列(autnmusly replicating DNA sequence,ARS)
(二)着丝粒DNA序列(centrmere DNA sequence, CEN)
(三)端粒DNA序列(telmere DNA sequence,TEL)
The Nbel Prize in Physilgy r Medicine 2009
"fr the discvery f hw chrmsmes are prtected by telmeres and the enzyme telmerase"
Fig. Telmere structure and micrscpic appearance
图9-26 端粒酶的作用示意图
端粒延长(ALT)的方式
Nature Reviews Mlecular Cell Bilgy 9, 177-182 (February 2008)
核型 (karytype)染色体组在有丝分裂中期的表型,包括染色体数目、大小、形态特征的总和核型模式图 (idigram)一个染色体组全部染色体逐个按其特征绘制下来, 再按长短、形态等特征排列起来的图象染色体显带技术经物理、化学因素处理后,再用染料对染色体进行分化染色,使其呈现特定的深浅不同带纹(band)的方法一类是产生的染色带分布在整过染色体长度上如:G、Q和R 带,一类是局部性显带,它只能使少数特定区域显带,如C、T和N 带
图9-27 人类细胞中期染色体显带及染色体大小示意图
用于核型分析的细胞培养
四、特殊染色体——多线染色体
存在于双翅目昆虫的幼虫组织细胞内来源于核内有丝分裂多线化的细胞处于永久间期多线染色体的带和间带都含有基因
图9-28 果蝇唾腺细胞全套多线染色体图中红色信号表示异染色质蛋白HP1 在多线染色体上的分布,绿色是dCAF-1-p180 蛋白的分布
四、特殊染色体——灯刷染色体
普遍存在于动物卵母细胞中,其中两栖类卵母细胞的灯刷染色体最典型卵母细胞进行减数分裂第一次分裂时停留在双线期的染色体;与卵子发生过程中营养物储备密切相关大部分DNA 以染色粒形式存在,没有转录活性侧环是RNA 活跃转录区域;合成的主要为前体mRNA
Figure 4-54 Mlecular Bilgy f the Cell (© Garland Science 2008)
Figure 4-55 Mlecular Bilgy f the Cell (© Garland Science 2008)
核仁(nuclelus)
真核细胞间期核中最显著的结构核仁的大小、形状和数目随生物的种类、细胞类型和细胞代谢状态不同而变化核仁是rRNA 合成、加工和核糖体亚单位的组装场所
Nucleli (nuclelus 的复数) HeLa cell, GFP-ribsmal prtein
纤维中心(fibrillar centers,FC):是被致密纤维包围的一个或几个低电子密度的圆形结构,主要成分为RNA聚合酶和rDNA,这些rDNA是裸露的分子致密纤维组分(dense fibrillar cmpnent,DFC):呈环形或半月形包围FC,由致密的纤维构成,是新合成的rRNA和特异性结合蛋白,转录主要发生在FC与DFC的交界处颗粒组分(granular cmpnent,GC):由直径15-20 nm的颗粒构成,是不同加工阶段的RNP
图9-29 BHK-21 细胞核仁的电镜照片银粒示rRNA 转录部位
核糖体的生物发生(ribsme bigenesis):包括rRNA 的合成、加工和核糖体亚单位的组装涉及mRNA 的输出与降解
三、核仁的动态周期变化
核仁周期(nuclelar cycle)核仁结构完整性(structure integrity)前核仁体 (prenuclelar bdy,PNB)核仁外体(extra nuclelar bdy)
四、核体(nuclear bdy)
亚核结构没有膜包被;高度动态变化包括:卡哈尔体(Cajal bdy)GEMS(Gemini f ciled bdy)染色质间颗粒(interchrmatin granule clusters, speckles)可能是蛋白质和 RNA 组分(或许也有DNA)相互作用的结果
Fig. Diversity f nuclear bdies
核基质 (nuclear matrix)
将DNA、组蛋白和RNA 抽提后,残留在核内的纤维蛋白网架结构,也称核骨架(nuclear skeletn)真核细胞核内的结构体系与核纤层、中间丝相互连接形成的网络体系,是贯穿于核与质的一个相对独立的结构系统主要成分由非组蛋白的纤维蛋白构成的,含有多种蛋白质成分。存在少量RNA 与DNA 复制、基因表达及染色体组装与构建密切相关
核被膜(nuclear envelpe)
位于细胞核的最外层,是细胞核与细胞质之间的界膜主要有3 种结构组分:双层核膜、核孔复合体与核纤层功能:构成了核、质之间的天然选择性屏障;调控细胞核内外的物质交换和信息交流
图9-1 细胞核截面图A. 细胞核结构组成示意图。B. 透射电镜图片显示胰腺细胞核:Nu,核仁;NPC,核孔复合体;Chr,染色质;NE,核被膜
由内外两层平行但不连续的单位膜构成内(层)核膜(inner nuclear membrane):无核糖体颗粒附着,紧贴其内表面有一层致密的纤维网络结构,即核纤层(nuclear lamina)外(层)核膜(uter nuclear membrane):常附有核糖体颗粒,与糙面内质网相连续核周间隙(perinuclear space)或核周池(perinuclear cisternae):20~40nm核孔(nuclear pre)核孔复合体(nuclear pre cmplex,NPC)孔膜区(pre membrane dmain)
一、核膜——崩解与组装
真核细胞中,核膜伴随着细胞周期的进行有规律地解体与重建核被膜去组装、重组装可能通过对核纤层蛋白、核孔复合体蛋白等进行磷酸化与去磷酸化修饰来实现
Nuclear envelpe breakdwn and reassembly in mitsis
核孔复合体是一个相对独立的复杂结构;间期细胞核普遍存在核孔复合体在核被膜上的数量、分布密度与分布形式随细胞类型、细胞核的功能状态的不同而有很大的差异研究核孔复合体形态结构的经典方法主要有三种:树脂包埋超薄切片技术、负染色技术、冷冻蚀刻技术
图9-2 用冷冻蚀刻电镜技术(A)和高分辨率扫描电镜(B)显示的核孔复合体(NPC)
胞质环(cytplasmic ring)核质环(nuclear ring)辐(spke):柱状亚单位;腔内亚单位;环带亚单位中央栓(central plug),中央颗粒 (central granule)
图9-3 核孔复合体结构模型
(一)结构模型—— “fish-trap”
Nature Reviews Mlecular Cell Bilgy 11, 490-501 (July 2010) | di:10.1038/nrm2928
(二)组成成分——核孔蛋白 (nucleprin,Nup)
gp210:结构性跨膜蛋白介导NPC与核被膜连接在内、外核膜融合形成核孔中起重要作用在核质交换中起作用p62:功能性Nup疏水性N 端区: 具有FXFG重复序列C 端区:具有疏水性的7肽重复序列
双功能、双向性的亲水性核质交换通道双功能表现在它有两种运输方式:被动扩散与主动运输双向性表现在既介导蛋白质的入核转运,又介导RNA、核糖核蛋白颗粒(RNP)的出核转运
1.通过核孔复合体的被动扩散
图9-4 通过核孔复合体物质运输的功能示意图
2.核孔复合体的主动运输
具有高度选择性对运输颗粒大小的限制是一个信号识别与载体介导的过程,需要消耗ATP 能量,并表现出饱和动力学特征双向性:核输入与核输出
亲核蛋白(karyphilic prtein)入核转运机制
亲核蛋白:在细胞质内合成后,需要或能够进入细胞核内发挥功能的一类蛋白质包括:在一个细胞周期中一次性地被转运到核内,并一直停留在核内行使功能的亲核蛋白,如组蛋白、核纤层蛋白等;需要穿梭于核质之间进行功能活动的亲核蛋白,如imprtin核定位序列(nuclear lcalizatin sequence)或核定位信号(nuclear lcalizatin signal,NLS):引导亲核蛋白入核,富含碱性氨基酸残基,如Lys、Arg,还常含有Pr;可以是一段连续的序列,也有分成两段存在;受体为imprtin;完成核输入后不被切除核输出信号(nuclear exprt signal,NES):引导RNP输出细胞核,受体为exprtinRan蛋白:属G蛋白,调节货物复合体的解体或形成
注射核质蛋白及核质蛋白组分的细胞定位
A classic experiment that used several frms f radiactive nucleplasmin(核质蛋白), which is a large pentameric prtein invlved in chrmatin assembly. Either the intact prtein r the nucleplasmin heads, tails, r heads with a single tail were injected int the cytplasm f a frg cyte r int the nucleus. All frms f nucleplasmin, except heads, accumulated in the nucleus when injected int the cytplasm, and all frms were retained in the nucleus when injected there.A. 核质蛋白哪部分负责其在核内定位?B.该实验如何证明其入核是由核定位信号指导的主动运输,而不是被动扩散?
核定位信号与Ran蛋白
亲核蛋白通过核孔复合体的转运:结合(binding)与转移(translcatin)
亲核蛋白NLS 识别imprtin α,与可溶性受体imprtin α/β 异二聚体结合,形成转运复合物在imprtin β 介导下,转运复合物与核孔复合体的胞质纤维结合转运复合物通过改变构象的核孔复合体从胞质面被转移到核质面转运复合物在核质面与Ran-GTP 结合,并导致复合物解离,亲核蛋白释放受体亚基与结合的Ran 返回胞质,在胞质内Ran-GTP 水解形成Ran-GDP 并与imprtin β 解离,Ran-GDP返回核内再转换成Ran-GTP 状态
图9-5 亲核蛋白从细胞质向细胞核输入的过程示意图
NPC介导双向核质转运
三、核纤层 nuclear lamina
3 种核纤层蛋白:lamin A、B 、C核纤层蛋白本身形成纤维状网络结构功能:结构支撑;调节基因表达;调节DNA 修复;与细胞周期有关
图9-6 冷冻蚀刻电镜技术显示核纤层的纤维结构
In HGPS patients the cell nucleus has dramatically aberrant mrphlgy (bttm, right) rather than the unifrm shape typically fund in healthy individuals (tp, right)
Hutchinsn-Gilfrd早衰综合征(HGPS):核纤肽A的C端缺失50个氨基酸
染色质 (chrmatin)
间期细胞核内由DNA、组蛋白、非组蛋白及少量RNA 组成的线性复合结构,是间期细胞遗传物质存在的形式细胞内基因存在与发挥功能的结构基础与基因组直接相关的细胞活动都是在染色质水平进行,如DNA 复制、基因转录、同源重组、DNA 修复,包括转录偶联的修复(transcriptin cupled repair)以及DNA 和组蛋白的各种修饰
图9-7 DNA 复制、转录和重组都是在染色质水平进行
一、染色质DNA——基因组大小比较
基因组(genme):某一生物的细胞中储存于单倍染色体组中的总遗传信息真核生物基因组DNA 的含量比原核生物高得多最小最简单的细胞支原体有迄今发现的能独立生活的有机体的最小基因组
Figure 1-37 Mlecular Bilgy f the Cell, Fifth Editin (© Garland Science 2008)
一、染色质DNA——基因组DNA 类型
蛋白编码序列编码rRNA、tRNA、snRNA和组蛋白的串联重复序列含有重复序列的DNA未分类的间隔DNA
DNA二级结构:两条多核苷酸链反向平行盘绕所生成的双螺旋结构分3 种:B 型DNA(右手双螺旋),“经典”的Watsn-Crick 结构;A 型DNA(B 型DNA 变构形式,右手双螺旋);Z 型DNA (B 型DNA 变构形式,左手螺旋)大沟的特征在遗传信息表达过程中起关键作用
组蛋白(histne)与DNA 结合没有序列特异性非组蛋白(nnhistne)与特定 DNA 序列或组蛋白相结合
属碱性蛋白质,富含带正电荷的Arg 和Lys 等碱性氨基酸,可以和酸性DNA 紧密结合,一般不要求特殊核苷酸序列核小体组蛋白 H2A、H2B、H3 和H4没有种属及组织特异性,进化上十分保守H1 组蛋白有一定的种属和组织特异性
(一)组蛋白(histne)
(二)非组蛋白(nnhistne)
序列特异性 DNA结合蛋白 特性多样性识别DNA具有特异性具有功能多样性
凝胶延滞实验(gel retardatin assay)
三、核小体(nuclesme)
核小体:染色质组装的基本结构单位染色质结构的“串珠”模型
图9-8 透射电镜显示串珠状11 nm 的核小体结构
透射电镜显示串珠状11 nm 的核小体结构
Figure 4-22 Mlecular Bilgy f the Cell (© Garland Science 2008)
盐溶液处理染色质11nm
非特异性微球菌核酸酶(micrcccal nuclease)消化染色质,DNA 被降解成约200 bp 的片段
X 射线晶体衍射揭示核小体三维结构
图9-9 由X 射线晶体衍射所揭示的人类细胞核小体三维结构A. 通过DNA 超螺旋中心轴所显示的核小体核心颗粒8 个组蛋白分子的位置。B. 垂直于中心轴的角度所见到的核小体核心颗粒的盘状结构
SV40 微小染色体(minichrmsme)分析
Virlgy, 2009, 384(2): 352-359
包括200 bp 左右的DNA 超螺旋和一个组蛋白八聚体以及一个分子的组蛋白H1组蛋白八聚体构成核小体的盘状核心颗粒,由4 个异二聚体组成,包括两 个H2A • H2B 和两个H3 • H4
146/147 bp:盘绕组蛋白八聚体1.75 圈166/167 bp:组蛋白H1 结合额外20 bp DNA,锁住核小体DNA的进出端,稳定核小体200 bpLinker DNA:相邻核小体之间;典型长度为60 bp,不同物种变化值为0~80 bp 不等
图 9-10 核小体的结构要点示意图
组蛋白与DNA 之间的相互作用主要是结构性的,基本不依赖于核苷酸的特异序列核小体沿DNA 的定位受不同因素的影响非组蛋白与DNA 特异性位点的结合,可影响邻近核小体的相位(psitining)富含AT 的DNA片段优先存在于DNA 双螺旋的小沟,而富含GC 的DNA 片段优先存在于DNA 双螺旋的大沟
Figure 4-23 Mlecular Bilgy f the Cell (© Garland Science 2008)
用稀酸或高盐溶液处理染色质,可使组蛋白与 DNA 解离,如何解释?
组蛋白与 DNA 之间的结合依靠的是组蛋白带正电的碱 性基团与 DNA 带负电荷的磷酸基团之间的静电引 力,如果用稀酸处理复合物,则磷酸基团质子化而 失去所带的负电荷,复合物解离。如果用高盐溶液 处理复合物,则阳离子与磷酸基团结合而取代了组 蛋白,导致组蛋白与 DNA 解离。
人体细胞46条染色体含DNA约6×109 bp,总长2米,如何计算出来的?
裸露DNA 组装直径30 nm 螺线管H3 • H4 两个异二聚体结合两个H2A • H2B 异二聚体加入组蛋白去乙酰化,H1组蛋白结合伴随着核小体折叠6 个核小体组成一个螺旋旋或由其他组装方式形成一个螺线管结构进一步折叠事件使染色质在核中形成确定的结构
图9-11 DNA 组装成染色质的过程及各阶段的协助组装因子
(一)染色质组装的前期过程
一级结构:核小体二级结构:螺线管三级结构:超螺线管四级结构:染色单体
图9-12 30 nm 染色质纤维的三维结构
(二)染色质组装的多级螺旋模型
(三)染色质组装的放射环结构模型
电镜:非组蛋白构成染色体骨架(chrmsmal scaffld)和与骨架相连的无数DNA 侧环染色体的骨架-放射环结构模型(scaffld radial lp structure mdel):30 nm 的染色线折叠成环,沿染色体纵轴,由中央向四周伸出,构成放射环
染色质具有高度复杂的结构
图9-13 基因组结构和染色质组装一览图
五、染色质类型 (一)常染色质与异染色质
常染色质(euchrmatin):间期细胞核内染色质纤维折叠压缩程度低,相对处于伸展状态,用碱性染料染色时着色浅异染色质 (heterchrmatin):间期细胞核中染色质纤维折叠压缩程度高,处于聚缩状态,用碱性染料染色时着色深结构异染色质或组成型异染色质(cnstitutive heterchrmatin):整个细胞周期均处于聚缩状态,没有较大变化;在中期染色体上多定位于着丝粒区、端粒、次缢痕等;由相对简单、高度重复的DNA 序列构成;具有显著的遗传惰性;为晚复制、早聚缩兼性异染色质 (facultative heterchrmatin):在某些细胞类型或一定的发育阶段,原来的常染色质聚缩,并丧失基因转录活性,变为异染色质;如巴氏小体(Barr bdy)
Barr Bdies: heterchrmatinized X-chrmsmes
异染色质或常染色质之间随着发育时期或细胞周期的变化可能相互转化异染色质与常染色质之间的转变常常需要伴随着一些组蛋白与DNA 修饰
图9-14 常染色质与异染色质转变过程中伴随的组蛋白修饰的变化
(二)常染色质与异染色质间的转变
活性染色质(active chrmatin)具有转录活性具有疏松的染色质结构非活性染色质(inactive chrmatin) 没有转录活性
(三)活性染色质与非活性染色质
1.活性染色质对DNase Ⅰ超敏感
活化染色质对DNaseⅠ的优先敏感性(preferential sensitivity)是可转录染色质一个基本特征活化染色质对DNaseⅠ的优先敏感性与HMG14 和HMG17两种非组蛋白有关,但不是唯一原因DNaseⅠ超敏感位点(DNaseⅠ hypersensitive site)的存在是活性染色质的特点
2.活性染色质的蛋白组成与修饰变化
活性染色质很少有组蛋白H1 与其结合;组蛋白乙酰化程度高,且H2B很少被磷酸化H2A 在活性染色质中很少有变异形式存在;H3 变种H3.3 只在活跃转录的染色质中出现HMG14 和HMG17 只存在于活性染色质中,与DNA 结合活性染色质标志:H3 N 端第4 个赖氨酸甲基化,第9 和14 个赖氨酸乙酰化以及第10 个丝氨酸磷酸化;非活性染色质标志:H3 N 端第9 个赖氨酸甲基化而不是乙酰化
图9-15 H3 组蛋白修饰与染色质活性的关系
染色质的三维动态分布与细胞ID
一、染色质的复制与修复
%20Glasgw%20June%202010/Image1.gif
S 期染色质完全复制不仅需要基因组DNA 的复制,也需要把复制好的DNA组装成染色质新复制的DNA 主要通过两种途径组装成染色质:第一,在复制叉的移动期间,父代的核小体核心颗粒与DNA 分离,到该段DNA 复制完成,父代的核小体核心颗粒直接转移到两条子链DNA 的一条上第二,染色质组装因子利用刚刚合成的、乙酰化的组蛋白介导核小体在复制DNA 上组装
图9-16 异染色质复制过程示意图
高度浓缩的染色质结构对基因组DNA(染色质DNA)具有重要的保护作用染色质DNA 随着细胞周期的变化会暴露或部分暴露于其周围环境中。基因组DNA 会受到细胞内外化学的与物理的因素的作用产生突变细胞中存在一整套的DNA 修复机器来应对各种各样的DNA 突变染色质修复(chrmatin restratin):修复后的DNA 组装成染色质的过程细胞一旦丧失了DNA 修复的部分或全部能力,基因组就会变得不稳定。基因组的不稳定性是直接导致肿瘤发生的重要原因之一
(二)染色质的修复与基因组稳定性
Fig. Chrmatin mdificatins and their cnsequences in respnse t lesins that are repaired by NER
二、染色质的激活与失活
染色质结构改变与基因活化的关系的研究主要集中在3 个方面:如何形成活性染色质,以便RNA 聚合酶能起始转录具有转录活性的染色质结构域如何与周围的非活性区域隔离RNA 聚合酶如何通过与染色质模板结合进行转录
染色质是一个动态、可塑的蛋白质与核酸组成的复合体调控蛋白结合到染色质DNA 的特定的位点上时,不管是在核小体间还是在一个核小体内,染色质都很容易被引发二级结构的改变。这些改变使得其他的一些结合位点与调控蛋白的结合变得要么更加容易,要么更加困难
图9-17 核小体变构因子通过改变核小体的相位而调节染色质的活性
(一)染色质的激活——DNA 结构与核小体相位的变化
核心组蛋白的赖氨酸残基乙酰化组蛋白H1 的磷酸化不同组蛋白修饰之间的关系
(一)染色质的激活——组蛋白的修饰
核心组蛋白的赖氨酸残基乙酰化(acetylatin)
图9-18 乙酰化和去乙酰化对染色质活性的影响A. 乙酰化和去乙酰化的位置。B. 导致染色质的变化
图9-19 酵母组蛋白乙酰化与去乙酰化控制转录的作用机制URS: 上游抑制序列(upstream repressin sequence);UAS: 上游激活序列;RD: 抑制结构域;AD: 激活结构域。
高速泳动族非组蛋白HMG1 和 HMG2 有A、B 和C 3 个结构域HMG 结构域蛋白结合在DNA 双螺旋的小沟中与DNA 弯折和DNA-蛋白质复合体高级结构形成有关
HMG (high-mbility grup)
(一)染色质的激活——HMG 蛋白的影响
图9-20 哺乳动物X 染色体失活模型Xic:X 染色体失活中心;Xist RNA:X 染色体失活特异性转录物;RNA(X-inactivatin specific transcript RNA)
(二)染色质的失活——X 染色体失活
活性基因转位到异染色质区附近时会失活的现象称为位置花斑效应(psitin effect variegatin)
(二)染色质的失活——位置花斑效应
三、染色质与基因表达调控
Circulatin Research. 2010; 107: 324-326
(一)以染色质为模板的转录
染色质是否处于活化状态(呈疏松或紧密结构)是决定RNA 聚合酶能否有效行使转录功能的关键转录起始伴随着染色质上某一基因调节序列内部或者周围的结构改变RNA聚合酶被认为是用“核小体犁”(nuclesme plw)来解除组蛋白和DNA 间的相互作用
图9-21 酵母激活因子及辅激活子有顺序地相互作用于HO 基因在染色质水平调节其表达
通用转录因子与结合RNA 聚合酶的核心启动子位点结合特异转录因子与特异基因的各种调控位点结合,促进或阻遏基因的转录DNA 结合域结合特异的DNA 序列激活结构域 激活转录
(二)转录因子介导的基因表达调控
转录抑制因子结合于特定的调节DNA 序列,抑制结合区附近基因转录DNA 结合域抑制结构域
图9-22 基因表达的时空特异性图中红色显示的是在果蝇的翅膀成虫盘(wing disc)中,dpp-GAL4 介导的Uif 的表达在前后分界处。绿色是在同一个盘中的背腹分界处的Wg 的表达。dpp:decapentaplegic;Uif:uninflatable;wg:Wingless
(三)DNA 甲基化介导的基因表达调控
DNA 甲基化(DNA methylatin)与基因表达的阻遏有关DNA 甲基化对维持基因长期处于非活化状态起重要作用。非活跃转录基因的DNA 甲基化程度普遍高于活跃转录的基因甲基化作用通过两种方式抑制转录:一是干扰转录因子对DNA 结合位点的识别,二是将转录激活因子识别的DNA 序列转换为转录抑制因子的结合位点DNA 甲基化在基因组印记(genmic imprinting)中具有重要作用
Fig. Regulatin f DNA methylatin f gene expressin
(四)组蛋白修饰介导的基因表达调控
转录因子能够调节组蛋白核心的结构,改变核小体和染色质的紧密程度,影响通用转录因子和RNA 聚合酶对启动子的结合,调控基因表达转录激活因子通常有利于那些导致染色质或组蛋白结构松散的蛋白质复合物发挥作用,如组蛋白乙酰化酶转录抑制因子通常会加强那些促进染色质结构紧密的蛋白质的作用,如组蛋白去乙酰化酶
( 五)染色质与表观遗传
表观遗传的变化是通过组蛋白和DNA 的不同修饰而实现的表观遗传修饰可以遗传但富于变化,通过改变染色质的结构而改变基因组信息的表达可改变染色质结构的途径主要包括:组蛋白密码子(histne cde)、染色质重塑(chrmatin remdeling)、组蛋白异型体(histne variants)、DNA 甲基化和非编码RNA(nncding RNAs)的作用
四、染色质的三维动态分布与细胞ID
染色质在细胞核内的分布不是均匀的,也不是静止不变的,而是存在时间和空间上的特异性特异分布的特性与细胞内的基因表达相关,更与细胞的状态直接相关细胞的身份(identity)有可能根据某些标志性DNA 序列的三维分布建立根类细胞ID 标准化信息库
Fig. Schematic representatin f hierarchical chrmatin rganizatin in plants
染色体是细胞在有丝分裂(或减数分裂)时遗传物质存在的特定形式,是间期细胞染色质结构紧密组装的结果由两条相同的姐妹染色单体(chrmatid)构成,彼此以着丝粒 (centrmere)相连根据着丝粒在染色体上所处的位置,分为4 种类型
图9-23 根据着丝粒位置进行的染色体分类图示
动粒结构域(kinetchre dmain) 内板(inner plate)中间间隙 (middle space)外板 (uter plate)纤维冠(fibruscrna)中央结构域(central dmain)配对结构域(pairing dmain)
图9-24 着丝粒的结构域组织
(二)染色体的其他主要结构
次缢痕(secndary cnstrictin)核仁组织区(nuclelar rganizing regin,NOR)随体(satellite)端粒(telmere)
二、染色体的功能元件(functinal element)
构成染色体DNA 的3 种关键序列(key sequence)称为染色体DNA 的功能元件DNA 复制起点着丝粒端粒“人造微小染色体”(artificial minichrmsme)
图9-25 真核细胞染色体的3 种功能元件示意图
(一)自主复制DNA序列(autnmusly replicating DNA sequence,ARS)
(二)着丝粒DNA序列(centrmere DNA sequence, CEN)
(三)端粒DNA序列(telmere DNA sequence,TEL)
The Nbel Prize in Physilgy r Medicine 2009
"fr the discvery f hw chrmsmes are prtected by telmeres and the enzyme telmerase"
Fig. Telmere structure and micrscpic appearance
图9-26 端粒酶的作用示意图
端粒延长(ALT)的方式
Nature Reviews Mlecular Cell Bilgy 9, 177-182 (February 2008)
核型 (karytype)染色体组在有丝分裂中期的表型,包括染色体数目、大小、形态特征的总和核型模式图 (idigram)一个染色体组全部染色体逐个按其特征绘制下来, 再按长短、形态等特征排列起来的图象染色体显带技术经物理、化学因素处理后,再用染料对染色体进行分化染色,使其呈现特定的深浅不同带纹(band)的方法一类是产生的染色带分布在整过染色体长度上如:G、Q和R 带,一类是局部性显带,它只能使少数特定区域显带,如C、T和N 带
图9-27 人类细胞中期染色体显带及染色体大小示意图
用于核型分析的细胞培养
四、特殊染色体——多线染色体
存在于双翅目昆虫的幼虫组织细胞内来源于核内有丝分裂多线化的细胞处于永久间期多线染色体的带和间带都含有基因
图9-28 果蝇唾腺细胞全套多线染色体图中红色信号表示异染色质蛋白HP1 在多线染色体上的分布,绿色是dCAF-1-p180 蛋白的分布
四、特殊染色体——灯刷染色体
普遍存在于动物卵母细胞中,其中两栖类卵母细胞的灯刷染色体最典型卵母细胞进行减数分裂第一次分裂时停留在双线期的染色体;与卵子发生过程中营养物储备密切相关大部分DNA 以染色粒形式存在,没有转录活性侧环是RNA 活跃转录区域;合成的主要为前体mRNA
Figure 4-54 Mlecular Bilgy f the Cell (© Garland Science 2008)
Figure 4-55 Mlecular Bilgy f the Cell (© Garland Science 2008)
核仁(nuclelus)
真核细胞间期核中最显著的结构核仁的大小、形状和数目随生物的种类、细胞类型和细胞代谢状态不同而变化核仁是rRNA 合成、加工和核糖体亚单位的组装场所
Nucleli (nuclelus 的复数) HeLa cell, GFP-ribsmal prtein
纤维中心(fibrillar centers,FC):是被致密纤维包围的一个或几个低电子密度的圆形结构,主要成分为RNA聚合酶和rDNA,这些rDNA是裸露的分子致密纤维组分(dense fibrillar cmpnent,DFC):呈环形或半月形包围FC,由致密的纤维构成,是新合成的rRNA和特异性结合蛋白,转录主要发生在FC与DFC的交界处颗粒组分(granular cmpnent,GC):由直径15-20 nm的颗粒构成,是不同加工阶段的RNP
图9-29 BHK-21 细胞核仁的电镜照片银粒示rRNA 转录部位
核糖体的生物发生(ribsme bigenesis):包括rRNA 的合成、加工和核糖体亚单位的组装涉及mRNA 的输出与降解
三、核仁的动态周期变化
核仁周期(nuclelar cycle)核仁结构完整性(structure integrity)前核仁体 (prenuclelar bdy,PNB)核仁外体(extra nuclelar bdy)
四、核体(nuclear bdy)
亚核结构没有膜包被;高度动态变化包括:卡哈尔体(Cajal bdy)GEMS(Gemini f ciled bdy)染色质间颗粒(interchrmatin granule clusters, speckles)可能是蛋白质和 RNA 组分(或许也有DNA)相互作用的结果
Fig. Diversity f nuclear bdies
核基质 (nuclear matrix)
将DNA、组蛋白和RNA 抽提后,残留在核内的纤维蛋白网架结构,也称核骨架(nuclear skeletn)真核细胞核内的结构体系与核纤层、中间丝相互连接形成的网络体系,是贯穿于核与质的一个相对独立的结构系统主要成分由非组蛋白的纤维蛋白构成的,含有多种蛋白质成分。存在少量RNA 与DNA 复制、基因表达及染色体组装与构建密切相关
相关课件
高中生物人教版 (2019)必修1《分子与细胞》第3节 细胞核的结构和功能集体备课课件ppt: 这是一份高中生物人教版 (2019)必修1《分子与细胞》第3节 细胞核的结构和功能集体备课课件ppt,共34页。PPT课件主要包含了教学目标,克隆探索的光辉岁月,细胞核,绝大部分有1个细胞核,细胞核的功能,实验设计是否严谨,核移植,黑美西螈细胞,白美西螈细胞,动态图等内容,欢迎下载使用。
2021学年第3节 细胞核的结构和功能课文内容课件ppt: 这是一份2021学年第3节 细胞核的结构和功能课文内容课件ppt,共11页。PPT课件主要包含了黑美西螈,白美西螈,卵细胞,胚胎细胞,去细胞核,黑色个体等内容,欢迎下载使用。
高中生物第3章 细胞的基本结构第3节 细胞核的结构和功能集体备课ppt课件: 这是一份高中生物第3章 细胞的基本结构第3节 细胞核的结构和功能集体备课ppt课件,共27页。PPT课件主要包含了细胞核的功能,变形虫,课堂练习1,细胞核的结构,染色质和染色体,技能训练,课堂练习2,学生自制模型等内容,欢迎下载使用。











